1. 引言
兔瘟是由兔瘟病毒(Rabbit hemorrhagic disease virus, RHDV)引起兔的一种急性、接触性传染病,以呼吸系统出血,实质器官水肿,淤血及出血变化为特征,该病传播快,发病急,发病率和死亡率极高,是影响兔养殖业发展的重要传染病 [1] 。目前该病广泛使用RHDV组织灭活苗来进行预防。随着分子生物学技术的发展,研究者逐渐将目光转向安全高效的基因工程疫苗的研制。已有报道兔瘟基因工程杆状病毒源亚单位疫苗已进入复合检验中,在新型疫苗的研制中取得突破性进展 [2] 。活载体疫苗是一种新型疫苗,利用基因工程方法改造病毒作为载体,能够表达特定保护性抗原或免疫调节因子等,特别是可以构建多价疫苗以达到一针防多病的目的 [3] 。疱疹病毒是一群较大的有囊膜的双链DNA病毒。I型牛疱疹病毒(Bovine hepervirus-1,BHV-1)为α疱疹病毒,具有疱疹病毒科成员所共有的形态特征。BHV-1宿主范围窄,不易传播于其他生物,尤其不会感染人类,因而将减少公共健康方面的忧虑。BHV-1基因组庞大,结构中有许多非必需基因,可以插入多个外源基因,并能使免疫动物产生外源表达产物的抗体,是成为构建多价疫苗的理想载体,现已有很多外源病毒DNA成功地插入到了BHV-1基因组中 [4] 。本研究以gE基因缺失BHV-1为活病毒载体,将兔瘟病毒唯一免疫原性基因VP60插入以获取重组病毒,能够在易感细胞稳定增殖和表达,为BHV-1作为病毒活载体研制兔瘟疫苗以及其他传染性疾病疫苗奠定了基础。
2. 材料和方法
2.1. 材料
牛肾细胞(MDBK),牛肺细胞(BL),gE基因缺失、并引入大肠杆菌β-半乳糖苷酶基因(LacZ)的重组病毒、含有BHV-1/gE 基因的上游同源重组序列及CMV控制下的LacZ基因的pGEMTgILdgELacZ质粒及含有BHV-1gE基因下游同源重组序列的质粒pGEM-TgESN均由哈尔滨兽医研究所大动物研究室构建并保存;含有RHDV VP60基因的重组质粒pMD-VP60由本哈尔滨兽医研究所自然疫源性人兽共患病研究室构建并保存。
2.2. 试剂
豚鼠抗RHDV VP60 单克隆抗体(MAb)由本实验室制备保存;辣根过氧化物酶标记和异硫氰酸荧光素标记的山羊抗鼠IgG、DMSO、X-gal购自Sigma公司;新生胎牛血清、MEM培养基、含0.25%胰蛋白酶和0.1%EDTA的细胞消化液为GIBCO产品;质粒提试剂盒、连接酶及内切酶均购自Promega公司;病毒DNA/RNA提取试剂盒购自天根生物科技有限公司;甲基纤维素(MC)、低熔点琼脂糖购自BBI公司;其他试剂为国产分析纯。
2.3. 细胞培养、病毒增殖和基因组提取
含10%新生胎牛血清的MEM用于培养MDBK和BT,用细胞消化液消化分散细胞并传代培养。待细胞长成单层后,接种无血清的病毒稀释液,37℃孵育1 h后,弃上层液体,补加含2%新生胎牛血清的MEM继续培养,当细胞病变达80%左右时回收细胞。4℃超声破碎细胞,2000 rpm低速离心5 min,回收上清。4℃ 10,000 rct高速离心30 min,弃上清留底部约200 μl,重复高速离心一次,最终留离心管底部100 μl液体用于病毒DNA提取。按病毒DNA/RNA提取试剂盒步骤提取病毒基因组用于PCR鉴定及转染试验。
2.4. 转移载体的构建
采用常规的分子生物学方法构建转移载体(如图1),将含有上游同源重组臂pGEM-TgILdgELacZ质粒与pMD-VP60质粒同时进行Hin d III和Xba I的双酶切,回收目的片段,纯化后连接构建重组质粒pGEMTgILdgEVP60;将其与含有下游同源重组臂pGEMTgESN质粒分别进行Sma I和Nsi I双酶切,回收目的片段纯化后进行连接构建含有目的基因表达盒的转移载体pGEM-TgILdgEVP60gESN重组质粒。该转移载体含有巨细胞病毒(cytomegalovirus immediate-early, CMV)早期启动子控制下的VP60基因以及用于同源重组的BoHV-1/gE基因上、下游同源重组序列。
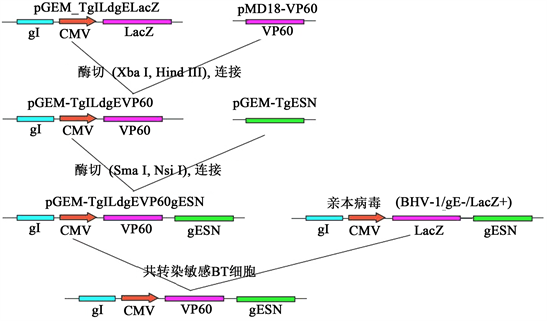
Figure 1. Construction of transfer vector and homologous recombination
图1. 转移载体的构建及同源重组
2.5. 转移载体与亲本病毒基因组的共转染
采用磷酸钙介导法进行转染,将2.5 μg经Ecot 22 I酶切的转移载体与亲本病毒基因组共转染BL细胞,以25%的DMSO休克3 min后,用无血清MEM洗涤两次,加入含10%胎牛血清MEM培养基,于37℃培养,待出现80%细胞病变(CPE)后收获培养物。
2.6. 重组病毒的筛选
将转染后收取的病毒液反复冻融后用0.22 μm的滤器过滤,10倍稀释后接种于MDBK细胞,37℃孵育1 h,弃去上清,然后覆盖以含2%血清和1%甲基纤维素的MEM,37℃培养,待出现明显CPE时弃去培养液并充分洗涤。然后无菌覆盖含3%血清和1%低熔点琼脂糖的MEM,并添加X-gal (150 μg/mL)进行染色。37℃培养24 h后观察蓝白斑情况,挑取白色病毒蚀斑继续进行蚀斑克隆纯化,直到所有病毒蚀斑均为白色。
2.7. VP60基因PCR鉴定
将筛选纯化病毒嗜斑接种MDBK细胞,待产生明显病变后收取病毒,提取病毒基因组。参照基因序列设计上游引物P1:5'-TGGATATCATGGAGGGCAAAGCCCG-3'和下游P2: 5'-CGGGATCCTCAGACATAAGAAAAGCCATTG-3',对VP60基因进行PCR鉴定。
2.8. Western Blot检测
重组病毒BHV-1-VP60及亲本病毒接种MDBK细胞,待出现明显细胞病变时收取细胞,超声破碎后用Millipore蛋白浓缩管浓缩后进行SDS-PAGE电泳,然后电转移至硝酸纤维素膜,以豚鼠抗RHDV VP60蛋白的MAb (1:100)为一抗,山羊抗鼠IgG-HRP (1:2000)为二抗,DAB显色检测蛋白表达情况。
2.9. 间接免疫荧光(IFA)检测
分别以100 TCID50的重组病毒及亲本病毒感染MDBK细胞,待CPE达80%时弃去培养液,以甲醇与丙酮按1:1混合固定细胞30 min,以豚鼠抗 RHDV VP60蛋白的MAb (1:50)为一抗,山羊抗鼠IgG-FITC (1:100)为二抗,荧光显微镜(Axiovert200)下观察,检测VP60蛋白表达情况。
3. 结果
3.1. 转移载体的构建及共转染
成功构建了含有VP60表达盒的转移载体pGEMTgILdgEVP1gESN,该转移载体含有用于同源重组的基因序列,经酶切及PCR鉴定,插入片段和扩增片断与理论大小一致(图2),测序结果正确;用该转移载体与亲本病毒的基因组共转染BL细胞后,可发生同源重组,经筛选后可获得重组病毒。
3.2. 重组病毒 BHV-1-VP60的筛选
依据LacZ在含有X-gal指示培养基上的显色反应,逆向筛选白色病毒嗜斑。经10轮的病毒蚀斑筛选后不再有蓝色病毒蚀斑出现(图3)。将白色病毒嗜斑连续纯化10轮后用于PCR及Western blot和间接免疫荧光试验检测。
3.3. 重组病毒BHV-1-VP60的PCR鉴定
将筛选的白色病毒蚀斑经MDBK细胞增殖后,提取病毒基因DNA,经VP60特异性引物扩增,琼脂糖凝胶电泳显示扩增出约1617 bp的特异性片段,与预期大小相同,而亲本病毒制备的DNA样品则
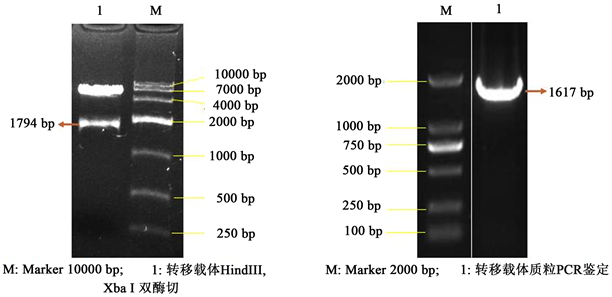
Figure 2. Identification of transfer vector for restriction enzyme digestion and PCR
图2. 转移载体的酶切和PCR鉴定
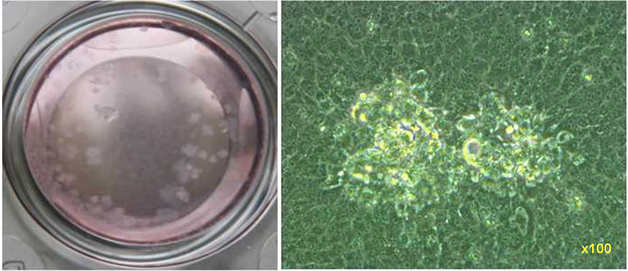
Figure 3. Observation of the recombinant virus
图3. 眼观及镜下观测所获重组病毒嗜斑
无特异性片段(图4)。所示结果表明RHDV VP60基因已经成功插入到重组病毒BHV-1-VP60基因组中,即已获得了在gE基因缺失位置插入外源基因的重组病毒。以噬斑纯化的重组病毒在MDBK细胞中连续传20代,取不同代次病毒 提取基因组DNA,经特异性引物扩增,结果显示均扩增出VP60基因,表明该重组病毒具有良好的遗传稳定性。
3.4. VP60基因表达的Western Blot检测
以重组病毒及亲本病毒感染MDBK细胞的裂解物进行SDS-PAGE,转移至硝酸纤维素膜后以抗VP60的单克隆抗体为一抗、HRP标记的山羊抗鼠IgG为二抗分别作用后于DAB中显色。Western blot结果显示在接种重组病毒BHV-1-VP60的样品中可见约60 kDa的免疫印迹条带,与预期VP60蛋白分子量值相符,VP60蛋白在MDBK细胞中获得了表达,而接种亲本病毒的样品则无免疫印迹条带(图5)。
3.5. VP60基因表达的IFA检测
IFA检测结果显示感染重组病毒BHV-1-VP60的BL细胞产生了红色的特异性荧光,VP60蛋白在MDBK细胞中获得了表达,而感染亲本病毒的BL细胞则无可见荧光(图6)。
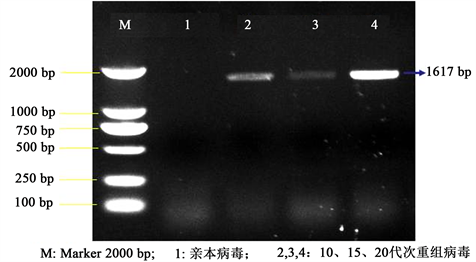
Figure 4. PCR identification for recombinant virus
图4. 重组病毒BHV-1-VP60的PCR鉴定
Figure 5. Western blot detection results of recombinant virus
图5. 重组病毒的Western blot检测结果
 (a)
(a)  (b)
(b)
Figure 6. Identification of recombinant virus by IFA. (a) Infected parent virus MDBK cells; (b) Infection recombinant virus MDBK cells
图6. 重组病毒BHV-1-VP60的IFA鉴定。(a) 感染亲本病毒MDBK细胞;(b) 感染重组病毒MDBK细胞
4. 讨论
RHD为上世纪末期暴发的一种危害养兔业的烈性传染病,针对RHD的预防只能通过注射疫苗来完成。目前国内研究学者已研制出多种常规疫苗,如RHD组织灭活苗,RHD油乳剂灭活苗,RHD氢氧化铝佐剂苗,RHD蜂胶佐剂灭活苗,热灭活苗等 [5] 。由于RHDV没有合适的体外繁殖系统,只能依赖传统方法从自然或人工感染致死的兔肝、肺组织中分离病毒抗原,不仅增加了实验的成本和周期,还给实验的准确性带来了很大的干扰。因此从基因工程学角度出发研制RHD新型疫苗势在必行。针对RHDV的唯一结构蛋白基因VP60能够在不同系统中表达的基因工程苗正在被研究开发,并已取得了较好的效果,如大肠杆菌表达亚单位疫苗,昆虫细胞表达亚单位疫苗,酵母细胞表达亚单位疫苗,马铃薯表达的亚单位疫苗等,以及重组牛痘病毒,金丝雀痘病毒,黏液病毒等活载体苗。活载体疫苗向宿主免疫系统提交免疫原性蛋白,可诱导产生体液免疫和细胞免疫,甚至黏膜免疫,同时兼有免疫效力高、成本低及安全性好等优点;若病毒载体中同时插入多个外源基因,就可以达到预防多种疾病的目的,因此活载体疫苗是当今与未来疫苗研制与开发的主要方向之一 [6] [7] 。随着疱疹病毒弱毒苗的问世及质量不断提高,以此为基础的活载体也逐渐成为研究目标。目前,疱疹病毒作为活载体表达外源基因的研究主要有:单纯疱疹病毒、伪狂犬病毒、火鸡疱疹病毒、牛疱疹病毒I型(BHV-1)、马疱疹病毒I型和传染性喉气管炎病毒等 [8] [9] 。本研究所采用BHV-1是目前从自然感染牛体内分离到的8种疱疹病毒之一,与其他疱疹病毒科的成员具有相同的形态,病毒核酸为线性双股DNA。牛是该病毒的主要宿主,而且BHV-1除了反刍动物之外,不存在其他保毒宿主,家兔可以通过鼻注或静注等途径感染而无发病症状 [10] ,而且在以BHV-1为活病毒载体的RHD基因工程疫苗的研究尚无相关报道。本研究是将中国兽医药品监察所于20世纪80年代从匈牙利引进的一株经过多年使用、证实安全性良好的BHV-1弱毒疫苗株BarthaNu/67人为缺失gE基因、并引入大肠杆菌β-半乳糖苷酶基因的重组病毒作为活载体,利用同源重组技术在病毒复制非必需区将VP60基因置于目前已知真核细胞中最强的启动子CMV之下,使得VP60基因能够更有效地表达;之后利用β-半乳糖苷酶基因在含X-gal培养基中特殊显色作用,逆向筛选表达VP60基因的白色病毒嗜斑进行纯化。Western blot和IFA检测结果表明VP60基因在MDBK细胞中得到了表达。重组病毒及亲本病毒在同一易感细胞中均能够产生CPE,这也表明VP60基因的插入并未影响重组病毒的感染性。
5. 结论
本实验成功构建表达RHDV-VP60基因的重组病毒BHV-1-VP60,且载体病毒背景明确。重组病毒BHV-1-VP60的构建将改变以往使用组织灭活苗和亚单位疫苗在免疫过程中的缺陷,具有安全、高效、操作简单、价格便宜等优点,有着良好的应用前景,为以后RHD活载体疫苗及多价疫苗的研制开发奠定了理论和实践基础。
基金项目
黑龙江省教育厅面上项目(12541580)。
NOTES
*通讯作者。