1. 引言
动物的能量平衡和调节机制是生理生态学的一个重要问题。自然环境中食物的质量和数量、环境温度、光周期等因素的季节行变化对动物的生存、繁殖和能量代谢等各方面都有重要影响[1] [2] 。20世纪80年代对褐色脂肪组织(brown adipose tissue, BAT)的研究促进了对体重调节机制的理解,Trier提出了食物诱导产热的概念[3] 。肥胖基因及其产物瘦素的研究,将体重调节机制的研究有推向了一个新的进展[4] 。Speakman等[5] 人提出了热散失限制理论,认为热量散失是能量稳态的主要限制因素。解偶联蛋白1 (uncoupling protein1, UCP1)是分布于线粒体内膜上的一种阴离子载体蛋白及产热蛋白[6] ,分子量为约32 KD,UCP1在产热调节中起关键作用[7] 。瘦素主要是由白色脂肪细胞分泌的、肥胖基因编码的、分子量为16 KD的蛋白类激素,在动物体重调节、发育于生殖、免疫等方面有重要作用[4] 。
中缅树鼩(Tupaia belangeri) 属于攀鼩目(Scadentia)树鼩科(Tupaiidae),是一类分布于南亚和东南亚、东洋界特有的小型哺乳动物,其主要分布与云南、广西、贵州、四川及海南岛等地,并且我国的云南、四川西南部和贵州很可能是树鼩分布的北限[8] 。树鼩由于具有特殊的进化地位,因此在生物医学上被广泛采用为实验动物。
2. 主要研究成果
本研究组自1990年以来,一直进行中缅树鼩的生理生态学研究,已经在中缅树鼩产热特征能量代谢各方面取得一系列研究成果:
2.1. 季节变化
限制能量消耗有三种假说,第一种是“中央限制”假说:限制能量消耗主要是中央能量消耗结构,当肠道的容纳和吸收能力最大时,动物的能量摄入受到限制;第二种是“外周限制”假说:当外周器官比如肌肉、褐色脂肪组织、乳腺等功能达到最大时,摄入能量后无法再利用,因此而受到限制;第三种假说是“热散失限制”假说:认为当热量散失达到最大时,为了防止身体过热,能量摄入受到限制[5] [9] 。
动物获得能量的多少决定了动物生长、繁殖和脂肪储存的能量,但是动物获得能量的速率和效率受到环境的限制,两个重要的环境因子是食物的供应和气候条件可能对动物能量收支的结构有直接的影响,因此,可以预测栖息地和季节的变化可能对野外的能量收支有重要的影响[10] 。动物对外部条件的变化做出回应的一种方式就是改变它们的消化特性,消化效率的可塑性被认为是对外部或是内部条件改变做出的最重要的生理适应[11] ,消化道作为能量摄入和能量分配之间的功能纽带,在生物进化过程中经历了许多选择压力[11] [12] 。消化道的形态学和生理学等可以决定动物对营养物质和能量摄入的吸收效率以及吸收速率[13] 。
从细胞、组织、器官以及整体等不同层次水平,整合分析了东洋界特有物种中缅树鼩在季节环境中的产热能力、能量代谢以及消化道研究发现:中缅树鼩在冬季来临时,靠增加体重、增加BMR和NST、增加能量摄入、消化能和可代谢能来对冬季环境变化做出回应,其代谢率和能量收支存在季节可塑性,并通过调节消化道长度和重量来满足能量需求的增加[14] ;同时,BAT总蛋白含量、线粒体蛋白含量以及细胞色素C氧化酶、α-磷酸甘油氧化酶活性,冬季也显著高于夏季,但增加的比例较温带物种低;而肝脏细胞总蛋白含量、线粒体蛋白含量以及细胞色素C氧化酶、α-磷酸甘油氧化酶活性以及线粒体呼吸状态3、状态4等,冬夏季节均无显著差异,因此,中缅树鼩产热能力介于热带和温带类群之间,显示出向温带类型过渡的趋势[15] 。
中缅树鼩的体重变化特征显著与北方高纬度地区小型啮齿动物不同,冬季体重显著高于夏季,很可能与中缅树鼩特殊分布区相关,中缅树鼩分布于与中南半岛相邻近的热带亚热带及其高原地区,与高纬度地区相比,冬季受低温胁迫要弱,即便是在冬季,食物资源仍然比较高纬度地区充足,因此,可以通过增加食物摄取来增加能量消耗,这可能是导致冬季体重高于夏季的原因之一,中缅树鼩冬季体重增加也可能导致体表面积相对减少,从而降低热丧失,有利于降低能量消耗,这一结果与热散失假说[16] 预计的结果相一致。中缅树鼩是热带亚热带种类,冷耐受能力相对较差,为了保持体温的恒定,中缅树鼩通过增加BMR来增加产热,从而能在气温较热带低的云南高原存活。中缅树鼩消化道长度和重量发生了变化,冬季较夏季增加,特别是小肠变化较大,这一结果与中央限制假说相一致,冬季摄入量的增加使得消化道增加容纳量,而夏季摄入量减少使得消化道容纳量增加受到限制[17] 。
2.2. 日节律变化
体温的日节律变化来源于两个方面,产热和散热,热的产生又来源于肌肉活动和非颤抖性生热作用[18] [19] 。经常讨论的一个因素就是活动性,在日节律中活动性的变化与体温节律有联系[20] [21] ,例如,金色仓鼠(Mesocricetus auratus)体温的变化有70%来源于活动性的变化[22] 。许多学者也研究发现动物体温和活动性的关系,发现活动性影响体温[23] ,例如,中缅树鼩的体温下午较高,清晨较低,表现出明显的日节律变化,且变化趋势与代谢率变化不同步,说明中缅树鼩体温的日节律变化可能与其下午活动性较高,而晚上和清晨活动性减少有关[24] 。中缅树鼩的BMR和NST也表现出日节律变化,它们的变化趋势与热中性区以下的代谢率变化趋势是同步的,在环境温度较低时代谢率较大,环境温度高时代谢率较小,说明中缅树鼩BMR和NST的日节律变化与环境温度的日节律变化有光[24] 。
2.3. 光照和温度对中缅树鼩能量代谢的影响
低温短光诱导中缅树鼩体重增加、体温降低、产热能力增强,静止代谢率增加的速率和比例高于非颤抖性产热,低温短光照刺激肝脏线粒体蛋白质量、线粒体氧化能力和细胞色素C氧化酶活性显著增加[25] ;短光照也刺激褐色脂肪组织总蛋白和线粒体蛋白量、线粒体状态4呼吸、细胞色素C氧化酶、α-磷酸甘油氧化酶及T4 5'-脱碘酶活性显著提高[26] 。低温和短光照促使血清T3浓度增加、T4浓度不变。中缅树鼩在低温短光照下体重增加,高温长光照下体重降低,与体重的季节性变化一致[27] -[29] 。短光照刺激中缅树鼩基础代谢率显著增加,与如欧鼠平[30] 、地中海田鼠[31] 等研究结果一致。
2.4. 冷暴露对褐色脂肪组织解偶联蛋白1、血清瘦素含量的影响
本研究组自备抗血清采用酶联免疫法测定了中缅树鼩在(5 ± 1)℃冷暴露0 d、7 d、14 d、21 d、28 d时,褐色脂肪组织(BAT)中解偶联蛋白1 (UCP1)的含量。结果表明,随着冷暴露时间的延长,中缅树鼩的体重、褐色脂肪组织重量均表现出了增加的趋势,BAT线粒体总蛋白和UCP1的含量也呈增加的趋势,其中UCP1的含量在28 d时达到极显著水平,比对照组增加了55.9%。说明冷暴露能够诱导中缅树鼩UCP1表达增加,从而使其适应性产热增加[32] 。
中缅树鼩的血清瘦素水平随着低温短光照时间的延长而显著降低,与体重呈显著负相关,其体重的调节方式与典型的北方小型兽类显著不同,表现出了一定的特异性,可能存在着瘦素抵抗的体重调节模式;胰岛素水平与瘦素水平呈显著的正相关,说明在中缅树鼩中,胰岛素能够正向的调节和刺激瘦素的分泌;低温短光照显著刺激了甲状腺素T3的增加,从而增强了产热能力,而高温长光照对其影响不大,但呈现出降低的趋势,在一定程度上减弱了产热的增加[25] 。冷暴露过程中肝脏线粒体蛋白含量对低温的反应比总蛋白的反应剧烈,心脏的线粒体蛋白含量也随着冷驯化时间的延长而显著增加,但腓肠肌的反应较为温和;各组织线粒体的状态IV呼吸能力均显著增强,除腓肠肌外其它各组织细胞色素C氧化酶活性也随冷暴露时间的延长而急剧增加[33] 。
2.5. 能量稳态的其他方面研究
注射外源性褪黑激素后,中缅树鼩体温明显降低,静止代谢率和非颤抖性产热显著增加,并随注射时间的延长增加的越多;注射褪黑激素不影响肝脏和褐色脂肪组织的重量,但两者线粒体蛋白含量显著增加、线粒体呼吸和细胞色素C氧化酶活性增加;褐色脂肪组织的α-磷酸甘油氧化酶和T4 5'-脱碘酶活性显著增强;血清T3浓度显著上升,T4浓度显著降低。结果表明,松果体褪黑激素参与了中缅树鼩的适应性产热调节[34] 。
中缅树鼩在夏季体温相对较高,受环境温度的影响较大;蒸发失水与环境温度显著正相关,在热中性区内基本维持相对稳定的水平,蒸发失水在体温调节中起着重要作用。结合同域分布的其他物种的生理生态学特征,提出中缅树鼩在热能代谢、体温调节及蒸发失水方面具有热带小型哺乳动物的特征,同时,又显示出某些适应于亚热带高原气候的特点[35] 。
2.6. 树鼩起源
目前关于现生树鼩的起源与分布,现在仍然尚不完全清楚,主要存在两种假说,即岛屿起源假说和大陆起源假说[36] 。从现生攀鼩目如印度树鼩(Anathana ellioti)、中缅树鼩、细尾树鼩(Dendrogale murina)和笔尾树鼩(Ptilocercus lowii)分布特征,强有力地支持树鼩起源于婆罗洲及其附近岛屿,即岛屿起源假说。除上述几种树鼩外,目前已经在泰国[37] 、印度[38] [39] 、巴基斯坦[40] (Jacobs, 1980)和中国[41] 等大陆的中新统地层和中国南部的始新世地层[42] 中发现几种树鼩的化石记录。值得注意的,最近在中国的元谋发现了细尾树鼩(或原细尾树鼩Prodendrogale)和笔尾树鼩的近亲[41] ,仅仅发现牙齿,并没有给出详细的描述和图片,若确实在中国南部中新世地层中存在树鼩这两种近亲[43] ,也将为大陆起源假说提供了支持。
如果树鼩起源于大陆,那么NSTmax对冷驯化中缅树鼩肝线粒体产热的贡献的变化趋势,应该与北方小型哺乳动物趋势相同,但中缅树鼩NST在冷适应产热中的比率逐渐降低,并且下降程度明显高于横断山固有种大绒鼠和由古北界迁移并现居于横断山地区的高山姬鼠,而横断山固有种大绒鼠下降程度明显高于由古北界迁移并现居于横断山地区的高山姬鼠,但本研究结果却刚好相反,NST在冷适应产热中的比率逐渐降低,因此中缅树鼩很可能是从热带迁移到现在的分布区,即由南向北扩散,这从生理生态方面为树鼩岛屿起源学说提供了一个依据[29] 。
经过我们实验室的多年研究发现,温度是限制中缅树鼩继续向北扩散的主要因素[29] ;中缅树鼩表现出不同于北方典型小型哺乳动物的产热特征:相对较为狭窄的热中性区,较高体温,代谢率较低;在冷驯化条件下,中缅树鼩体重增加,基础代谢率在总产热所占比例高于非颤抖性产热,血清瘦素含量与体重呈负相关,UCP 1增加的程度相对较低。
3. 研究展望
关于小型哺乳动物能量代谢和稳态调节可能是表型及其可塑性研究最多的领域之一,也是研究表型及其可塑性变化对生存适应和自然选择最适合的领域[44] 。动物的生存必须获取和同化能量,并且将同化的能量按需要分配到维持生存[9] [45] 、生长和繁殖[46] 等各方面,而且通过个体的生理过程与种群的生态模式紧密地联系起来[47] (图1)。
近几年来,关于脂肪组织的研究取得了突破性进展,其中最重要的突破是发现决定BAT细胞形成肌源性起源机制,从而改变了过去许多关于BAT细胞形成及其功能的认识:如重新发现并证实了健康成年人体内依然存在具有功能的BAT细胞[48] -[50] ,并且人体和实验小鼠体内的BAT细胞与WAT细胞相混合[51] [52] ,冷暴露[49] 季节变化[53] 有显著的适应性反应。在实验室小鼠体内也证实了这一现象[54] 。确定了BAT细胞的分化起源主要为肌源性起源[55] 。从而为深入研究能量稳态维持、能量平衡及其表型可塑性变化提供了新思路[56] [57] 。目前认为BAT细胞肌源性起源主要包括两条途径(图2),即PRDM16和骨骼形成蛋白7 (BMP7)途径。
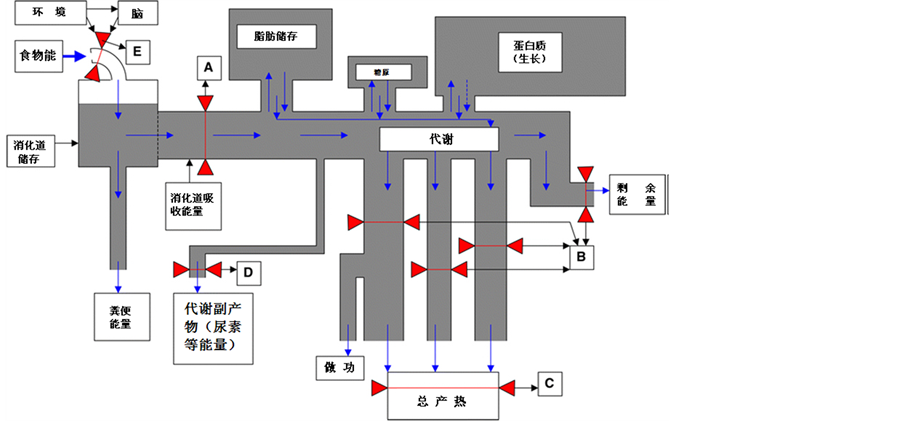
Figure 1. Schematic diagram to show energy flows in an animal and the potential points at which such energy flows may be constrained and thus control the level of the sustained energy intake. A—Central limitation hypothesis; B—Peripheral limitation hypothesis; C—Heat dissipation limit theory; D—Metabolic end products discharge limits; E—Affect food intake through adjustment mechanism of compensation in the brain. Blue arrows in figure indicated energy flow direction, red triangle indicated potential limit position.
图1. 能量在动物体内流动以及可能限制能量流动的部位,这些限制部位很可能是限制持续摄入能水平的部位;A——“中央”限制假说;B——外周限制假说;C——热散失限制假说;D——代谢尾产物排泄的限制;E——通过脑的补偿调节机制影响摄食量。图中蓝色箭头表示能流方向,红色三角形表示可能限制部位[9]
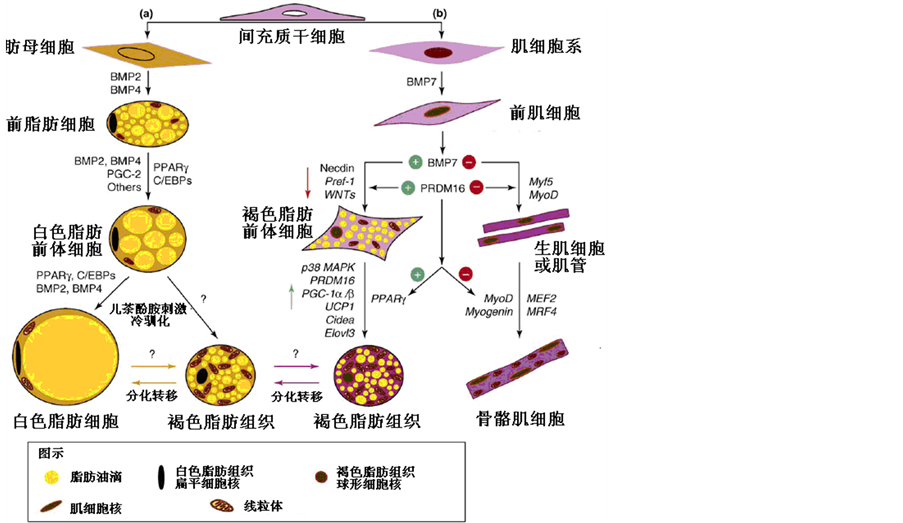
Figure 2. Mesenchymal stem cells differentiate into fat cells and muscle cells
图2. 间质干细胞分化成脂肪细胞和肌细胞[58]
“脂肪自稳理论”(lipostatic theory)假设“体脂信号”(adiposity signal)激素决定哺乳动物能量稳态维持以来,瘦素(Leptin)的发现起到划时代的意义。但是,Leptin刺激厌食症、增强分解的功能并不一定完全表现出来。尽管循环血液中Leptin水平与体脂储存之间存在一定的关系,但是,不仅肥胖症患者而且在啮齿动物中与Leptin水平增加相联系的并不一定表现出抑制食欲增强,这种现象称之为Leptin抵抗,通常认为是扰乱有机体能量稳态维持的关键因素[4] 。Leptin抵抗主要表现出病理性,例如食物诱导性肥胖,或为允许体重调节点出现变化的适应性反应,例如哺乳动物的妊娠或动物的季节性变化[5] 。
有关中缅树鼩能量稳态以及褐色脂肪组织的肌源性起源的相关研究,我们实验室将在今后进行深入研究,探讨。
致谢
感谢国家国际科技合作专项项目(2014DFR31040);十二五科技支撑项目(2014BAI01B00);国家自然科学基金项目(No.31360096;No.31260097);云南省应用基础研究计划重点项目(No. 2013FA014)的支持。

NOTES
*第一作者。
#通讯作者。